Provirophages and transpovirons as the diverse mobilome of giant viruses
October 16, 2012 3:55 PM Subscribe
Provirophages and transpovirons as the diverse mobilome of giant viruses
(*#*) indicates that the article referenced is openly accessible
Introduction:
Abstract: A distinct class of infectious agents, the virophages1 that infect giant viruses of the Mimiviridae family, has been recently described. Here we report the simultaneous discovery of a giant virus of Acanthamoeba polyphaga (Lentille virus) that contains an integrated genome2 of a virophage (Sputnik 2), and a member of a previously unknown class of mobile genetic elements3, the transpovirons4. The transpovirons are linear DNA elements of ∼7 kb [kilobases]5 that encompass six to eight protein-coding genes, two of which are homologous6 to virophage genes. Fluorescence7 in situ hybridization8 showed that the free form of the transpoviron replicates within the giant virus factory and accumulates in high copy numbers inside giant virus particles, Sputnik 2 particles, and amoeba cytoplasm. Analysis of deep-sequencing data showed that the virophage and the transpoviron can integrate9 in nearly any place in the chromosome of the giant virus host and that, although less frequently, the transpoviron can also be linked to the virophage chromosome. In addition, integrated fragments of transpoviron DNA were detected in several giant virus and Sputnik genomes. Analysis of 19 Mimivirus strains revealed three distinct transpovirons associated with three subgroups of Mimiviruses. The virophage, the transpoviron, and the previously identified self-splicing introns10 and inteins11 constitute the complex, interconnected mobilome12 of the giant viruses and are likely to substantially contribute to interviral gene transfer.[Full Text PDF] and two explanations in English
(*#*) indicates that the article referenced is openly accessible
Introduction:
Mobile genetic elements (MGEs) that are collectively referred to as the “mobilome” are key players in the genome evolution of prokaryotes (*1*) and eukaryotes (*2*, *3*) and are considered “genetic engineers” of biological innovation (*1*). MGEs can be roughly grouped into four major classes: transposable elements (TEs), plasmids, viruses, and self-splicing elements such as group I and II introns and inteins (*4*). The mobilomes of many bacteria, archaea, and unicellular eukaryotes include all of these elements in a free or integrated form. Given that viruses constitute a part of the mobilome, they are not normally considered to possess mobilomes of their own. However, some large viruses contain retrovirus sequences integrated into their genomes (*5*, *6*), whereas others, including members of the Mimiviridae family, harbor self-splicing introns and/or inteins (*7*, *8*, *9*). Furthermore, many viruses support the reproduction of satellite viruses1 (*10*). The discovery of the Sputnik virophage in 2008 added a new twist to the existing understanding of the relationships between different mobile elements by demonstrating for the first time that a giant virus could be infected by another, much smaller virus in a manner similar to the viral infection of cells (11). The Sputnik virophage is a small icosahedral virus (74 nm in diameter) that parasitizes on Mamavirus, a member of the Mimiviridae family (12, 13). Sputnik replicates inside Mamavirus or Mimivirus viral factories when the host giant virus is grown in amoebae such as Acanthamoeba castellanii or A. polyphaga (11). An in-depth analysis of the Sputnik proteins has suggested an evolutionary connection between this virophage and a distinct class of TEs (14). The second virophage, the Mavirus (15), was isolated as a parasite of a distinct member of the Mimiviridae family, Cafeteria roenbergensis virus (CroV) [Previously on Metafilter] (16). At least four Mavirus proteins, including the major capsid protein13, are homologous6 to proteins of Sputnik. In addition, the Mavirus genome encodes a retroviral-type integrase and a protein-primed DNA polymerase B; these proteins are homologous to the respective proteins of Maverick/polinton DNA transposons, which insert into genomes of diverse eukaryotes, suggesting an evolutionary link between the Mavirus and the polintons (15). The third complete virophage genome sequence has been identified in the metagenome of the hypersaline Organic Lake in Antarctica (*17*). This Organic Lake virophage (OLV) is thought to parasitize on phycoDNAviruses that infect green algae. The OLV genome encodes seven proteins with homologs in Sputnik (*17*), including two key proteins, the major capsid protein and the DNA-packaging ATPase, that are shared by all three virophages. Thus, the virophages apparently share a common origin, although each underwent multiple gene replacements. The virophages are likely to be common parasites of nucleocytoplasmic large DNA viruses that infect diverse eukaryotes, and show multiple evolutionary connections to other mobile elements (*18*). Here we present findings that substantially expand the complexity of the giant virus mobilome through the description of an integrated form of the virophage and of a distinct class of MGEs, the transposovirons.Discussion:
The discovery of the Mimivirus and subsequent identification of other giant viruses revealed unexpected complexity of viral genomes that, with over 1,000 protein-coding genes, are more complex than many parasitic and symbiotic bacteria and are comparable to the most compact genomes of free-living bacteria and archaea (*7*). The present work shows that giant viruses are associated with a commensurately complex mobilome that encompasses three of the four major classes of mobile elements, namely self-splicing elements, transposable elements or linear plasmids (transpovirons), and viruses (virophages that can form provirophages after integration into the host giant virus genome). Different components of the giant virus mobilome share homologous genes, and genomic comparisons point to DNA transfer between the mobilome components and the host virus but also within the mobilome itself. Thus, the giant virus mobilome is a network that potentially could provide routes and vehicles for gene exchange and might make substantial contributions to the shaping of mosaic viral genomes. The giant viruses and their mobilomes together are part of even more expansive, dynamic genetic networks: the amoebae with their diverse bacterial parasites and symbionts and their own viruses (30).Glossary of Terms Used in the Abstract, Introduction, and Discussion:
Of special note is the transpoviron, a distinct plasmid that depends on giant viruses for its replication and spread. Substantial analogies can be found between the transpovirons and virus-associated plasmids present in bacteria and archaea. In particular, the well-studied bacteriophage P4 (also known as a “phasmid”) is a plasmid that replicates episomally in the absence of the helper bacteriophage P2 but is encapsidated into virions and thus can infect new bacterial cells in the presence of the helper (*31*, *32*). A similar replication strategy has been described for the archaeal virus plasmid pSSVx that depends on the fuselloviruses SSV1 or SSV2 and appears to have acquired genes from a fusellovirus (33). The discovery of the transpoviron shows that virus-associated plasmids exist in all three domains of cellular life.
It is unlikely that the present study exhausts the diversity of the giant virus mobilome; additional virophages and transpovirons, and perhaps distinct classes of mobile elements, are likely to be discovered. Indeed, the transpoviron had not been detected until the isolation of Lentille virus from a human sample described here. Furthermore, we failed to detect closely related homologs of transpoviron genes in the available databases of environmental sequences, although close homologs of many Mimivirus and Sputnik genes were readily detectable (11). Thus, specific conditions and/or habitats could be required for accumulation of transpovirons and probably other elements comprising the giant virus mobilome. Characterization of such conditions will likely lead to the discovery of additional genetic elements associated with giant viruses and facilitate elucidation of their replication mechanisms and the relationships between different mobilome components.
1Virophage: A subviral agent composed of nucleic acid that depends on the co-infection of a host cell with a helper or master virus for its multiplication. When a satellite encodes the coat protein in which its nucleic acid is encapsidated it is referred to as a satellite virus. A satellite virus of mimivirus that inhibits the replication of its host has been termed a virophage. However, the usage of this term remains controversial due to the lack of fundamental differences between virophages and classical satellite viruses.References from the Introduction and Discussion:
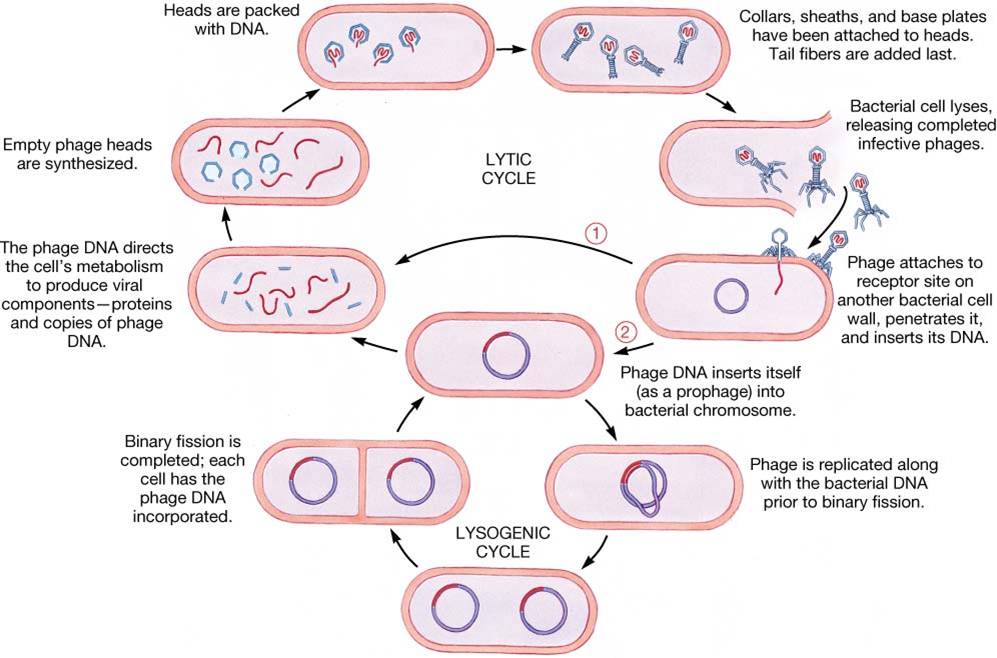
2Integrated Genome: A prophage is a phage (viral) genome inserted and integrated into the circular bacterial DNA chromosome. A prophage, also known as a temperate phage, is any virus in the lysogenic cycle; it is integrated into the host chromosome or exists as an extrachromosomal plasmid. Technically, a virus may be called a prophage only while the viral DNA remains incorporated in the host DNA. This is a latent form of a bacteriophage, in which the viral genes are incorporated into the bacterial chromosome without causing disruption of the bacterial cell. Upon detection of host cell damage, such as UV light or certain chemicals, the prophage is excised from the bacterial chromosome in a process called prophage induction. After induction, viral replication begins via the lytic cycle. In the lytic cycle, the virus commandeers the cell's reproductive machinery. The cell may fill with new viruses until it lyses or bursts, or it may release the new viruses one at a time in a reverse endocytotic process. The period from infection to lysis is termed the latent period. A virus following a lytic cycle is called a virulent virus. Prophages are important agents of horizontal gene transfer, and are considered part of the mobilome.
3Mobile genetic elements: Mobile genetic elements (MGE) are a type of DNA that can move around within the genome. They include Transposons (also called transposable elements including Retrotransposons, DNA transposons, and Insertion sequences), Plasmids, Bacteriophage elements like Mu (which integrates randomly into the genome), and Group II introns. The total of all mobile genetic elements in a genome may be referred to as the mobilome.
4Transpoviron: This is a new term that the authors are proposing to refer to mobile genetic elements within viruses.
5Kb or Kilobasepairs: A measurement of the length of a stretch of DNA equivalent to 1,000 base pairs
6Homology: Homologous traits of organisms are due to sharing a common ancestor, and such traits often have similar embryological origins and development. This is contrasted with analogous traits: similarities between organisms that were not present in the last common ancestor of the taxa being considered but rather evolved separately. An example of analogous traits would be the wings of bats and birds, which evolved separately but both of which evolved from the vertebrate forelimb and therefore have similar early embryology. Whether or not a trait is homologous depends on both the taxonomic and anatomical levels at which the trait is examined. For example, the bird and bat wings are homologous as forearms in tetrapods. However, they are not homologous as wings, because the organ served as a forearm (not a wing) in the last common ancestor of tetrapods. By definition, any homologous trait defines a clade—a monophyletic taxon in which all the members have the trait (or have lost it secondarily); and all non-members lack it
7Fluorescence: The emission of light by a substance that has absorbed light or other electromagnetic radiation. It is a form of luminescence. In most cases, the emitted light has a longer wavelength, and therefore lower energy, than the absorbed radiation. However, when the absorbed electromagnetic radiation is intense, it is possible for one electron to absorb two photons; this two-photon absorption can lead to emission of radiation having a shorter wavelength than the absorbed radiation. The emitted radiation may also be of the same wavelength as the absorbed radiation, termed "resonance fluorescence".
8In situ hybridization: A type of hybridization that uses a labeled complementary DNA or RNA strand (i.e., probe) to localize a specific DNA or RNA sequence in a portion or section of tissue (in situ)
9Viral Integration: A superpower possessed by Eukaryotic retroviruses (like HIV and Human T-lymphotropic virus) and temperate bacteriophages, where the virus integrates its genome into its host's genome and shuts off ll of its host lethal genes in order to hide - indefinitely - until conditions become better for an active infection.
10Intron: An intron is any nucleotide sequence within a gene that is removed by RNA splicing while the final mature RNA product of a gene is being generated. The term intron refers to both the DNA sequence within a gene, and the corresponding sequence in RNA transcripts. Sequences that are joined together in the final mature RNA after RNA splicing are exons. Introns are found in the genes of most organisms and many viruses, and can be located in a wide range of genes, including those that generate proteins, ribosomal RNA (rRNA), and transfer RNA (tRNA). When proteins are generated from intron-containing genes, RNA splicing takes place as part of the RNA processing pathway that follows transcription and precedes translation. The word intron is derived from the term intragenic region; i.e., a region inside a gene. Although introns are sometimes called intervening sequences, the term "intervening sequence" can refer to any of several families of internal nucleic acid sequences that are not present in the final gene product, including inteins, untranslated sequences (UTR), and nucleotides removed by RNA editing, in addition to introns.
11Intein: An intein is a segment of a protein that is able to excise itself and rejoin the remaining portions (the exteins) with a peptide bond. Inteins have also been called "protein introns".
12Mobilome: The total of all mobile genetic elements in a genome; a play on the word Genome.
13Major Capsid Protein: A capsid is the protein shell of a virus. It consists of several oligomeric structural subunits made of protein called protomers. The observable 3-dimensional morphological subunits, which may or may not correspond to individual proteins, are called capsomeres. The capsid encloses the genetic material of the virus.
1. Frost LS, Leplae R, Summers AO, Toussaint A (2005) Mobile genetic elements: The agents of open source evolution. Nat Rev Microbiol 3(9):722–732.Bonus:
2. Feschotte C, Pritham EJ (2007) DNA transposons and the evolution of eukaryotic genomes. Annu Rev Genet 41:331–368.
3. Kazazian HH, Jr. (2004) Mobile elements: Drivers of genome evolution. Science 303 (5664):1626–1632.
4. Siefert JL (2009) Defining the mobilome. Methods Mol Biol 532:13–27.
5. Sun AJ, et al. (2010) Functional evaluation of the role of reticuloendotheliosis virus long terminal repeat (LTR) integrated into the genome of a field strain of Marek’s disease virus. Virology 397(2):270–276.
6. Hertig C, Coupar BEH, Gould AR, Boyle DB (1997) Field and vaccine strains of fowlpox virus carry integrated sequences from the avian retrovirus, reticuloendotheliosis virus. Virology 235(2):367–376.
7. Raoult D, et al. (2004) The 1.2-megabase genome sequence of Mimivirus. Science 306 (5700):1344–1350.
8. Colson P, et al. (2011) Viruses with more than 1,000 genes: Mamavirus, a new Acanthamoeba polyphaga mimivirus strain, and reannotation of Mimivirus genes. Genome Biol Evol 3:737–742.
9. Van Etten JL (2003) Unusual life style of giant chlorella viruses. Annu Rev Genet 37: 153–195.
10. Simon AE, Roossinck MJ, Havelda Z (2004) Plant virus satellite and defective interfering RNAs: New paradigms for a new century. Annu Rev Phytopathol 42: 415–437.
11. La Scola B, et al. (2008) The virophage as a unique parasite of the giant mimivirus. Nature 455(7209):100–104.
12. Sun S, et al. (2010) Structural studies of the Sputnik virophage. J Virol 84(2):894–897.
13. Desnues C, Raoult D (2010) Inside the lifestyle of the virophage. Intervirology 53(5): 293–303.
14. Iyer LM, Abhiman S, Aravind L (2008) A new family of polymerases related to superfamily A DNA polymerases and T7-like DNA-dependent RNA polymerases. Biol Direct 3:39.
15. Fischer MG, Suttle CA (2011) A virophage at the origin of large DNA transposons. Science 332(6026):231–234.
16. Fischer MG, Allen MJ, Wilson WH, Suttle CA (2010) Giant virus with a remarkable complement of genes infects marine zooplankton. Proc Natl Acad Sci USA 107(45): 19508–19513.
17. Yau S, et al. (2011) Virophage control of Antarctic algal host-virus dynamics. Proc Natl Acad Sci USA 108(15):6163–6168.
18. Koonin EV, Yutin N (2010) Origin and evolution of eukaryotic large nucleo-cytoplasmic DNA viruses. Intervirology 53(5):284–292.
30. Raoult D, Boyer M (2010) Amoebae as genitors and reservoirs of giant viruses. In- tervirology 53(5):321–329.
31. Briani F, Dehò G, Forti F, Ghisotti D (2001) The plasmid status of satellite bacterio- phage P4. Plasmid 45(1):1–17.
32. Lindqvist BH, Dehò G, Calendar R (1993) Mechanisms of genome propagation and helper exploitation by satellite phage P4. Microbiol Rev 57(3):683–702.
33. Arnold HP, et al. (1999) The genetic element pSSVx of the extremely thermophilic crenarchaeon Sulfolobus is a hybrid between a plasmid and a virus. Mol Microbiol 34 (2):217–226.
Carl Zimmer's explanation of the debate over whether these Nucleocytoplasmic Large DNA Viruses (Giant) viruses constitute a new fourth domain of life.
If this whole post seems horribly intimidating starting out I would recommend beginning with one of the two excellent articles on Not Exactly Rocket Science or The Scientist linked above the fold.
If you now have a burning desire to restart you biology education from scratch, your friendly neighborhood microbial virologist recommends the biology classes offered for free on the MIT Open Courseware
posted by Blasdelb at 4:09 PM on October 16, 2012 [4 favorites]
If you now have a burning desire to restart you biology education from scratch, your friendly neighborhood microbial virologist recommends the biology classes offered for free on the MIT Open Courseware
posted by Blasdelb at 4:09 PM on October 16, 2012 [4 favorites]
Self-replicating molecules gonna self-replicate.
posted by lalochezia at 4:16 PM on October 16, 2012 [1 favorite]
posted by lalochezia at 4:16 PM on October 16, 2012 [1 favorite]
Wait, are we submitting posts for peer review now?
Damn impressive post Blasdelb.
posted by leotrotsky at 4:19 PM on October 16, 2012 [9 favorites]
Damn impressive post Blasdelb.
posted by leotrotsky at 4:19 PM on October 16, 2012 [9 favorites]
Is every living thing in the world bleeding genetic material?
posted by Nomyte at 4:20 PM on October 16, 2012
posted by Nomyte at 4:20 PM on October 16, 2012
Nomyte: "Is every living thing in the world bleeding genetic material?"
If something is you'll probably find their stuff in a Bdelloid Rotifer
posted by Blasdelb at 4:26 PM on October 16, 2012 [1 favorite]
If something is you'll probably find their stuff in a Bdelloid Rotifer
posted by Blasdelb at 4:26 PM on October 16, 2012 [1 favorite]
I must admit some disappointment that "giant viruses" do not obligate me to carry a machete at all times.
posted by CynicalKnight at 4:40 PM on October 16, 2012 [3 favorites]
posted by CynicalKnight at 4:40 PM on October 16, 2012 [3 favorites]
Blasdelb I'm really glad you did this.
posted by Buckt at 4:50 PM on October 16, 2012 [2 favorites]
posted by Buckt at 4:50 PM on October 16, 2012 [2 favorites]
leotrotsky: "Wait, are we submitting posts for peer review now?"
Reviewer Comment: This post contains insufficient cowbell and should be summarily dismissed
Response: We agree that, historically, there has been a great need for additional cowbell in MetaFilter posts, however, this post both acknowledges the great contributions that cowbell has provided for MeFite studies and is entirely orthogonal to it.
Reviewer Comment: This post is a SLYT, all purveyors of SLYT posts should be dragged out and shot (Actual phrase I've seen used in a reviewer comment - the reviewer was upset about the wide angle magnification of a TEM micrograph having failed to understand the purpose of the micrograph)
Response: We agree with the criticism and have added fluff on lines 4 through 118
posted by Blasdelb at 4:52 PM on October 16, 2012 [10 favorites]
Reviewer Comment: This post contains insufficient cowbell and should be summarily dismissed
Response: We agree that, historically, there has been a great need for additional cowbell in MetaFilter posts, however, this post both acknowledges the great contributions that cowbell has provided for MeFite studies and is entirely orthogonal to it.
Reviewer Comment: This post is a SLYT, all purveyors of SLYT posts should be dragged out and shot (Actual phrase I've seen used in a reviewer comment - the reviewer was upset about the wide angle magnification of a TEM micrograph having failed to understand the purpose of the micrograph)
Response: We agree with the criticism and have added fluff on lines 4 through 118
posted by Blasdelb at 4:52 PM on October 16, 2012 [10 favorites]
If something is you'll probably find their stuff in a Bdelloid Rotifer
That abstract introduced me to the wonderful word spliceosome. Now I'm sorry I didn't study microbiology, but a good 30% of my school's undergrad population were boring, disaffected, pre-med bio majors.
posted by Nomyte at 5:11 PM on October 16, 2012
That abstract introduced me to the wonderful word spliceosome. Now I'm sorry I didn't study microbiology, but a good 30% of my school's undergrad population were boring, disaffected, pre-med bio majors.
posted by Nomyte at 5:11 PM on October 16, 2012
I'm gonna favorite this. and then attempt to and probably fail at understanding it.
posted by ninjew at 5:36 PM on October 16, 2012
posted by ninjew at 5:36 PM on October 16, 2012
I think this post is playing host to several other posts which are themselves infected by other posts...
(I mean that in a good way)
posted by vorpal bunny at 5:41 PM on October 16, 2012 [4 favorites]
(I mean that in a good way)
posted by vorpal bunny at 5:41 PM on October 16, 2012 [4 favorites]
This is great, Blasdelb. Love all your posts about virology. Virophages/provirophages are new to me.
Also, amoebas are insane things.
posted by zennie at 5:42 PM on October 16, 2012 [2 favorites]
Also, amoebas are insane things.
posted by zennie at 5:42 PM on October 16, 2012 [2 favorites]
If virii have taught be anything, it's that life is information. And information will, given time, find a way to express itself.
posted by SPrintF at 6:30 PM on October 16, 2012 [2 favorites]
posted by SPrintF at 6:30 PM on October 16, 2012 [2 favorites]
Wow, how I have not heard of these?! "Yo dawg, I heard you like virii..."
posted by exogenous at 6:52 PM on October 16, 2012
posted by exogenous at 6:52 PM on October 16, 2012
This is some good stuff!! Many thanks for the 'translation' pages to ease me into the juicy stuff.
Bdelloid Rotifer? Those girlz are wicked!
It's not the giant viruses that scare me, it's the little evil deadly ones.
How come those little virusites get their own mobile 'ome? What's wrong with a tent? Sheesh, phages and virons today have it so easy.
posted by BlueHorse at 7:29 PM on October 16, 2012 [1 favorite]
Bdelloid Rotifer? Those girlz are wicked!
It's not the giant viruses that scare me, it's the little evil deadly ones.
How come those little virusites get their own mobile 'ome? What's wrong with a tent? Sheesh, phages and virons today have it so easy.
posted by BlueHorse at 7:29 PM on October 16, 2012 [1 favorite]
Big fleas have little fleas,
Upon their backs to bite 'em,
And little fleas have lesser fleas,
and so, ad infinitum.
posted by awenner at 7:58 PM on October 16, 2012 [1 favorite]
Upon their backs to bite 'em,
And little fleas have lesser fleas,
and so, ad infinitum.
posted by awenner at 7:58 PM on October 16, 2012 [1 favorite]
I love the absolute genetic promiscuity of viruses. You want some DNA? Not same species? No problem, take and do what you can with it. This must be where evolution boils the hottest.
posted by CautionToTheWind at 1:37 AM on October 17, 2012
posted by CautionToTheWind at 1:37 AM on October 17, 2012
"This must be where evolution boils the hottest."
If there are 1031 bacteriophages on the planet and they each have a half life about one day, that means there are 1031 phages made every two days, which means 6 x 1025 new phage particles produced per second on Earth. Even if we are really conservative and guess that only around one in every 10 million of those phages has a novel and beneficial mutation or recombination, that is still 6 x 1018, or six quintillion, completely new phages born every second. This is what one quintillion pennies looks like.
posted by Blasdelb at 5:35 AM on October 17, 2012 [6 favorites]
If there are 1031 bacteriophages on the planet and they each have a half life about one day, that means there are 1031 phages made every two days, which means 6 x 1025 new phage particles produced per second on Earth. Even if we are really conservative and guess that only around one in every 10 million of those phages has a novel and beneficial mutation or recombination, that is still 6 x 1018, or six quintillion, completely new phages born every second. This is what one quintillion pennies looks like.
posted by Blasdelb at 5:35 AM on October 17, 2012 [6 favorites]
Too many notes.
posted by Goofyy at 7:53 AM on October 17, 2012 [1 favorite]
posted by Goofyy at 7:53 AM on October 17, 2012 [1 favorite]
Too many notes
That's what they said about Beethoven, but he showed them!
This FPP prompted an obtuse question:
Would it be at all feasible to engineer (or conceivably, discover in the wild) a virophage or bacteriophage that could be used therapeutically? Is it possible there's some phage that's highly pathogenic to our pathogens, and could be used to help kill off (for example) AIDS in an infected human?
posted by Sleeper at 9:53 AM on October 17, 2012
That's what they said about Beethoven, but he showed them!
This FPP prompted an obtuse question:
Would it be at all feasible to engineer (or conceivably, discover in the wild) a virophage or bacteriophage that could be used therapeutically? Is it possible there's some phage that's highly pathogenic to our pathogens, and could be used to help kill off (for example) AIDS in an infected human?
posted by Sleeper at 9:53 AM on October 17, 2012
About the same time I posted that I discovered the Wikipedia article "phage therapy":
http://en.wikipedia.org/wiki/Phage_therapy
Looks like it's an idea that's only a little older than the discovery of phages themselves, but hasn't yet produced much in the way of useful results, in part because little research has been devoted to it.
I'm curious about the danger of creating a "there was an old lady who swallowed a fly" situation with phage therapy.
posted by Sleeper at 10:09 AM on October 17, 2012
http://en.wikipedia.org/wiki/Phage_therapy
Looks like it's an idea that's only a little older than the discovery of phages themselves, but hasn't yet produced much in the way of useful results, in part because little research has been devoted to it.
I'm curious about the danger of creating a "there was an old lady who swallowed a fly" situation with phage therapy.
posted by Sleeper at 10:09 AM on October 17, 2012
"Would it be at all feasible to engineer (or conceivably, discover in the wild) a virophage or bacteriophage that could be used therapeutically? Is it possible there's some phage that's highly pathogenic to our pathogens, and could be used to help kill off (for example) AIDS in an infected human?"
Oh Jesus Christ, I am a phage biologist who needs to go to bed.
Phages are just about the most awesome things ever but they can't infect AIDS, which doesn't have any known satellite viruses. They can however infect bacterial pathogens:
They are the viruses of bacteria, think herpes, influenza, smallpox, or HIV but for bacteria. This is what they look like, this is what they do, and to really make sense of them the story starts with an aggressively stubborn French-Canadian microbiologist named Félix d'Herelle.
After he and his brother lost family's fortune on a doomed chocolate factory, he left for South America where he made a decent living inventing new processes for converting tropical plants such as bananas and sisal into distilled liquor for western markets. However, while he was in Mexico he noticed something interesting, after the the swarms of locusts that devastated local agriculture passed through, sick locusts could be noticed to have been left behind. It occurred to him to isolate the pathogen to see if he could use it to combat the swarms. His technique ended up working so well that in 1911 d'Herelle was invited to travel to Argentina as a microbiologist to address the locust problems there.
The problem was MASSIVE, every other year locusts would create the modern equivalent of billions of dollars worth of damage to cash crops and generate famine on the extraordinarily fertile pampas. It was so bad, and Argentina was rich enough then, that plans were being drawn up to import most of the world’s silver to build massively long 4 meter high walls across the pampas to stop the plagues. Apparently they'd have done it to if they thought they could stop theft, and on top of this the Ministry of Agricultural Defense had grown to a 3,000 member strong bureaucracy dedicated to extraordinary campaigns to defeat them.
d'Herelle's plan was again to spread diseases of the locust itself ahead of the swarms to use the same terrifying scale that made farmers so helpless, against the plague. He ended up getting funding to find sick locusts, cultivate the disease though serial transfer between 100 locust cages, and thus isolate 100% virulent and contagious strains of a cocobacillus. When thousands of these carcasses were spread out ahead of a swarm they were brought to an epic halt within a few days. After two years of d'Herelle's efforts the plagues ceased to be the issue that they once were in Argentina and the Pasteur Institute sent out his cultures to Columbia where several successful trials were conducted, as well as Cyprus and Algeria where they had significant effect.
All of this must have primed him to the idea that pathogens, or at least problem species, might have their own pathogens that we could team up with for productive effects, thus perhaps contributing to one of the most brilliant examples of deductive reasoning in modern science. At one point while he was in Paris before 1917, d'Herelle noticed something odd in a lawn of dysentery bacteria he had grown on a petri dish, a glassy clear dead spot. He must have thought this was interesting and so he plucked the spot from the plate and spread it out over a new lawn of the same bacteria, which then would not grow. Presumably figuring that he had isolated a new toxin of some kind, he made serial dilutions of it to see how just how toxic it was, and it did something toxins had never been known to do before. Arranging the plates in a row from highest dilution to lowest dilution, for a toxin, one would expect to see progressively but evenly damaged growth as one went down the series. However he saw first low numbers and then high numbers of the same glassy spots that mathematically followed the series. He quickly made a leap of judgment that would be challenged by many of the finest minds in the word until he was proven right by one of the first electron micrographs ever taken, that this wasn't a toxin at all, but a discrete organism. The problem was that his phages were far too small to see with a light microscope, no matter how powerful, as visible light has a wavelength of around 600nm and phage are around 25-250nm (thus using light to get a sense of what phage look like is kind of like using the blunt end of a telephone pole to get a sense of what a grasshopper feels like).
While d'Herelle continued to work with phages, which I will get back to later, the next big advances in understanding what phages really are would wait almost a decade for a mass movement of out of work physicists who, having suddenly run out of things to do when we figured out to much of physics, came to biology the 1920s to the 1930s. They brought with them a mechanistic view of how the universe works that they used to cause massive transformations in how we understand and interact with biology, and most used phages. One of the most influential of these scientific interlopers was a charismatic guy named Max Delbrück who quickly reasoned that, if we were ever going to understand how life works, we would need to start with the simplest organism possible and work our way up. He isolated seven bacteriophages against E. coli B, originally just his lab strain, and named them in a series T1 (previously) through T7. The central idea was that he and his growing number of colleagues1 would focus on truly understanding how these phages worked and use that knowledge to generalize to Escherichia coli, then the mouse, and then us. An essential component of this was the "Phage Treaty" among researchers in the field, which Delbrück organized in order to limit the number of model phage and hosts so that folks could meaningfully compare results. What came out of their original focus on these phages, in many respects encapsulated in Erwin Schrödinger's What is life?, has shed light on so much as to truly redefine our self-understanding as a species, much less medicine:
His discovery of phages was long before antibiotics, when bacterial disease killed almost everyone eventually and in horrific ways without much anyone could do for the sick. d'Herelle instantly saw the value that this pathogen of bacteria could have for patients, just like the value his coccobacilli had for farmers. He soon found a chicken farm with chicken typhoid that he successfully treated with phage isolated from the farm itself. He then isolated bacteria from the stool of a bunch dying French cavalrymen at a military hospital, isolated phage against them, amplified those phages, purified them as best he could, drank a bunch to demonstrate safety, and then gave it to the cavalrymen who each very quickly recovered.
Phage therapy exploded quickly, the major pharmaceutical companies of the United States and Europe, including Eli Lily which is still around, pumped out cocktails as quickly as they could and marketed them aggressively. However, no one really knew what phages were, much less how they worked, and most of the commercial entities profiting from phage didn’t seem to much care. This ended up giving phage a very well deserved bad reputation among physicians who tried preparations that we now know to have been heat or acid killed, or against the wrong pathogen, or against the right pathogen but with the wrong host range, or advertised as being effective against absurd things like gallstones and herpes and understandably decided the whole thing was bullshit. Many physicians considered the question settled with a pretty damning article series published in JAMA in 19342, before antibiotics became available a few years later making the question at least seem largely irrelevant for most pathogens (Though successful phage therapy of typhoid fever continued in the 50s when effective antibiotics were finally found against S. typhii, and in France until the 80’s when poorly worded AIDS related legislation killed it).
Phage therapy did, however, survive and thrive in the Soviet Union after Stalin ended up reading d’Herelle’s first two books in the early 1930s with great interest. In 1934 he invited d’Herelle to set up a phage institute in what is now the Republic of Georgia with a Georgian microbiologist, George Eliava, for the purpose of studying phage and providing the Red Army with a reliable supply. While d’Herelle is said to have been initially enamored with communism, he was soon soured on it when Eliava was suddenly kidnapped, murdered, and denounced by Beria (it likely had as much to do with Beria demonstrating that even Heroes of Soviet Science were not immune to his power as anything else, but the oral history remembered by Georgian phage biologists is that Eliava slept with an opera singer that Beria had his eye on). Despite the institute’s decapitation with the loss of Eliava and the fleeing of d’Herelle, the women they trained took over and turned it into one of the great centers of Soviet medicine. They conducted large and well-designed, particularly for the era, studies to establish phage as a standard of care and then slowly expanded that standard as new needs arose.
Over the last fifteen years or so, with the breakup of the Soviet Union and the exponentially growing crisis of antibiotic resistance, phage therapy is looking very exciting again. Unlike the ‘30s, we now have a decent understanding of phage biology as well as the infrastructure to keep phages cold until use, effective diagnostic tools, and most importantly, regulatory structures that shut out hucksters. The need is also dire, for example multi-drug resistant Staph infections kill more people in the United States than AIDS does.
Basic phage biology has also been undergoing a resurgence as we discover just how important phages are to the global ecosystem, they are indeed the dominant organism on Earth outnumbering anything else by two orders of magnitude. Indeed, despite being just ~125 nm tall (check this out for scale), if one were to stack the 1031 phages on the planet end to end you would get a tower that would stand 200 million lightyears tall. Our oceans are remarkably free of cellular life and the reason is phages, as well as the other the viruses of microbes. For example the growth and death of algae blooms are centrally mediated by viral dynamics. Meta-genomics studies of the oceans pull out more predicted phage proteins than anything else without some fancy filtering, and even then they get a bunch. Phages are also teaching us a lot about the primordial origins of life, they are after all proto-cellular organisms.
These days are exciting times to be interested in phages.
Further reading:
Abedon S., Kuhl S., Blasdel B., & Kutter B. Phage Treatment of Human Infections. Bacteriophage 2011; 1:66–85 (PDF)
Krisch HM, Comeau AM. The immense journey of bacteriophage T4--from d'Hérelle to Delbrück and then to Darwin and beyond. Res Microbiol. 2008 Jun;159(5):314-24. Epub 2008 Jun 25.
Fruciano DE. Phage as an antimicrobial agent: d’Herelle's heretical theories and their role in the decline of phage prophylaxis in the West. Can J Infect Dis Med Microbiol. 2007 January; 18(1): 19–26.
Chanishvili N. Phage Therapy-History from Twort and d'Herelle Through Soviet Experience to Current Approaches. Adv Virus Res. 2012;83:3-40.
1Frank Stahl famously wrote: "The Phage Church, as we were sometimes called, was led by the Trinity of Delbrück, Luria, and Hershey. Delbrück's status as founder and his ex-cathedra manner made him the pope, of course, and Luria was the hard-working, socially sensitive priest-confessor. And Al (Hershey) was the saint."
2Eaton MD, Stanhope BJ. Bacteriophage therapy. JAMA 1934;103:1769-76.
posted by Blasdelb at 3:40 PM on October 17, 2012 [9 favorites]
Oh Jesus Christ, I am a phage biologist who needs to go to bed.
Phages are just about the most awesome things ever but they can't infect AIDS, which doesn't have any known satellite viruses. They can however infect bacterial pathogens:
They are the viruses of bacteria, think herpes, influenza, smallpox, or HIV but for bacteria. This is what they look like, this is what they do, and to really make sense of them the story starts with an aggressively stubborn French-Canadian microbiologist named Félix d'Herelle.
{kind=link}
After he and his brother lost family's fortune on a doomed chocolate factory, he left for South America where he made a decent living inventing new processes for converting tropical plants such as bananas and sisal into distilled liquor for western markets. However, while he was in Mexico he noticed something interesting, after the the swarms of locusts that devastated local agriculture passed through, sick locusts could be noticed to have been left behind. It occurred to him to isolate the pathogen to see if he could use it to combat the swarms. His technique ended up working so well that in 1911 d'Herelle was invited to travel to Argentina as a microbiologist to address the locust problems there.
The problem was MASSIVE, every other year locusts would create the modern equivalent of billions of dollars worth of damage to cash crops and generate famine on the extraordinarily fertile pampas. It was so bad, and Argentina was rich enough then, that plans were being drawn up to import most of the world’s silver to build massively long 4 meter high walls across the pampas to stop the plagues. Apparently they'd have done it to if they thought they could stop theft, and on top of this the Ministry of Agricultural Defense had grown to a 3,000 member strong bureaucracy dedicated to extraordinary campaigns to defeat them.
d'Herelle's plan was again to spread diseases of the locust itself ahead of the swarms to use the same terrifying scale that made farmers so helpless, against the plague. He ended up getting funding to find sick locusts, cultivate the disease though serial transfer between 100 locust cages, and thus isolate 100% virulent and contagious strains of a cocobacillus. When thousands of these carcasses were spread out ahead of a swarm they were brought to an epic halt within a few days. After two years of d'Herelle's efforts the plagues ceased to be the issue that they once were in Argentina and the Pasteur Institute sent out his cultures to Columbia where several successful trials were conducted, as well as Cyprus and Algeria where they had significant effect.
All of this must have primed him to the idea that pathogens, or at least problem species, might have their own pathogens that we could team up with for productive effects, thus perhaps contributing to one of the most brilliant examples of deductive reasoning in modern science. At one point while he was in Paris before 1917, d'Herelle noticed something odd in a lawn of dysentery bacteria he had grown on a petri dish, a glassy clear dead spot. He must have thought this was interesting and so he plucked the spot from the plate and spread it out over a new lawn of the same bacteria, which then would not grow. Presumably figuring that he had isolated a new toxin of some kind, he made serial dilutions of it to see how just how toxic it was, and it did something toxins had never been known to do before. Arranging the plates in a row from highest dilution to lowest dilution, for a toxin, one would expect to see progressively but evenly damaged growth as one went down the series. However he saw first low numbers and then high numbers of the same glassy spots that mathematically followed the series. He quickly made a leap of judgment that would be challenged by many of the finest minds in the word until he was proven right by one of the first electron micrographs ever taken, that this wasn't a toxin at all, but a discrete organism. The problem was that his phages were far too small to see with a light microscope, no matter how powerful, as visible light has a wavelength of around 600nm and phage are around 25-250nm (thus using light to get a sense of what phage look like is kind of like using the blunt end of a telephone pole to get a sense of what a grasshopper feels like).
While d'Herelle continued to work with phages, which I will get back to later, the next big advances in understanding what phages really are would wait almost a decade for a mass movement of out of work physicists who, having suddenly run out of things to do when we figured out to much of physics, came to biology the 1920s to the 1930s. They brought with them a mechanistic view of how the universe works that they used to cause massive transformations in how we understand and interact with biology, and most used phages. One of the most influential of these scientific interlopers was a charismatic guy named Max Delbrück who quickly reasoned that, if we were ever going to understand how life works, we would need to start with the simplest organism possible and work our way up. He isolated seven bacteriophages against E. coli B, originally just his lab strain, and named them in a series T1 (previously) through T7. The central idea was that he and his growing number of colleagues1 would focus on truly understanding how these phages worked and use that knowledge to generalize to Escherichia coli, then the mouse, and then us. An essential component of this was the "Phage Treaty" among researchers in the field, which Delbrück organized in order to limit the number of model phage and hosts so that folks could meaningfully compare results. What came out of their original focus on these phages, in many respects encapsulated in Erwin Schrödinger's What is life?, has shed light on so much as to truly redefine our self-understanding as a species, much less medicine:
Delbrück turned out to be absolutely right to start simple, and his branch of Biophysics turned into molecular genetics (as opposed to the Drosophila variety) and split off into modern genetics, molecular biology, protein biology, molecular physiology, bioengineering, as well as genomics and the various other –omics. It all started with phage, but around the 70s phage biology did start to die as old professors dies and retired while their students became leaders in all of these new and exciting fields. However, I promised I’d get back to d’Herelle.The Luria–Delbrück experiment elegantly demonstrated that in bacteria genetic mutations arise in the absence of selection, rather than being a response to selection. Evolutionary biology has made so much more sense ever since.
The Hershey–Chase experiment showed once and for all that nucleic acids were in fact the heritable molecule.
The two guys who discovered the model for the structure of the B form double helix were phage folk. Incidentally they published it in easily the snarkiest, most badass, and likely most important published scientific paper ever, written as an accessible single page. The structure of DNA, and its relationship to function that they discovered, is true for all of life.
Most of the central dogma, was also figured out using phage, from most of the functions of RNA to the triplicate nature of codons
So many of the enzymes, molecular tools, we now take for granted come from phage
His discovery of phages was long before antibiotics, when bacterial disease killed almost everyone eventually and in horrific ways without much anyone could do for the sick. d'Herelle instantly saw the value that this pathogen of bacteria could have for patients, just like the value his coccobacilli had for farmers. He soon found a chicken farm with chicken typhoid that he successfully treated with phage isolated from the farm itself. He then isolated bacteria from the stool of a bunch dying French cavalrymen at a military hospital, isolated phage against them, amplified those phages, purified them as best he could, drank a bunch to demonstrate safety, and then gave it to the cavalrymen who each very quickly recovered.
Phage therapy exploded quickly, the major pharmaceutical companies of the United States and Europe, including Eli Lily which is still around, pumped out cocktails as quickly as they could and marketed them aggressively. However, no one really knew what phages were, much less how they worked, and most of the commercial entities profiting from phage didn’t seem to much care. This ended up giving phage a very well deserved bad reputation among physicians who tried preparations that we now know to have been heat or acid killed, or against the wrong pathogen, or against the right pathogen but with the wrong host range, or advertised as being effective against absurd things like gallstones and herpes and understandably decided the whole thing was bullshit. Many physicians considered the question settled with a pretty damning article series published in JAMA in 19342, before antibiotics became available a few years later making the question at least seem largely irrelevant for most pathogens (Though successful phage therapy of typhoid fever continued in the 50s when effective antibiotics were finally found against S. typhii, and in France until the 80’s when poorly worded AIDS related legislation killed it).
Phage therapy did, however, survive and thrive in the Soviet Union after Stalin ended up reading d’Herelle’s first two books in the early 1930s with great interest. In 1934 he invited d’Herelle to set up a phage institute in what is now the Republic of Georgia with a Georgian microbiologist, George Eliava, for the purpose of studying phage and providing the Red Army with a reliable supply. While d’Herelle is said to have been initially enamored with communism, he was soon soured on it when Eliava was suddenly kidnapped, murdered, and denounced by Beria (it likely had as much to do with Beria demonstrating that even Heroes of Soviet Science were not immune to his power as anything else, but the oral history remembered by Georgian phage biologists is that Eliava slept with an opera singer that Beria had his eye on). Despite the institute’s decapitation with the loss of Eliava and the fleeing of d’Herelle, the women they trained took over and turned it into one of the great centers of Soviet medicine. They conducted large and well-designed, particularly for the era, studies to establish phage as a standard of care and then slowly expanded that standard as new needs arose.
Over the last fifteen years or so, with the breakup of the Soviet Union and the exponentially growing crisis of antibiotic resistance, phage therapy is looking very exciting again. Unlike the ‘30s, we now have a decent understanding of phage biology as well as the infrastructure to keep phages cold until use, effective diagnostic tools, and most importantly, regulatory structures that shut out hucksters. The need is also dire, for example multi-drug resistant Staph infections kill more people in the United States than AIDS does.
Basic phage biology has also been undergoing a resurgence as we discover just how important phages are to the global ecosystem, they are indeed the dominant organism on Earth outnumbering anything else by two orders of magnitude. Indeed, despite being just ~125 nm tall (check this out for scale), if one were to stack the 1031 phages on the planet end to end you would get a tower that would stand 200 million lightyears tall. Our oceans are remarkably free of cellular life and the reason is phages, as well as the other the viruses of microbes. For example the growth and death of algae blooms are centrally mediated by viral dynamics. Meta-genomics studies of the oceans pull out more predicted phage proteins than anything else without some fancy filtering, and even then they get a bunch. Phages are also teaching us a lot about the primordial origins of life, they are after all proto-cellular organisms.
These days are exciting times to be interested in phages.
Further reading:
Abedon S., Kuhl S., Blasdel B., & Kutter B. Phage Treatment of Human Infections. Bacteriophage 2011; 1:66–85 (PDF)
Krisch HM, Comeau AM. The immense journey of bacteriophage T4--from d'Hérelle to Delbrück and then to Darwin and beyond. Res Microbiol. 2008 Jun;159(5):314-24. Epub 2008 Jun 25.
Fruciano DE. Phage as an antimicrobial agent: d’Herelle's heretical theories and their role in the decline of phage prophylaxis in the West. Can J Infect Dis Med Microbiol. 2007 January; 18(1): 19–26.
Chanishvili N. Phage Therapy-History from Twort and d'Herelle Through Soviet Experience to Current Approaches. Adv Virus Res. 2012;83:3-40.
1Frank Stahl famously wrote: "The Phage Church, as we were sometimes called, was led by the Trinity of Delbrück, Luria, and Hershey. Delbrück's status as founder and his ex-cathedra manner made him the pope, of course, and Luria was the hard-working, socially sensitive priest-confessor. And Al (Hershey) was the saint."
2Eaton MD, Stanhope BJ. Bacteriophage therapy. JAMA 1934;103:1769-76.
posted by Blasdelb at 3:40 PM on October 17, 2012 [9 favorites]
A couple of dumb questions about phage(s): 1. Do you pronounce it fayj or fahj? and 2. Is phage singular or plural? (Or is it short for "phage therapy"?)
I've had these questions ever since I heard a Science Friday on phage therapy. The Russian (I think) gentleman pronounced it fahj, and used it like a plural, e.g. "this is what phage can do". I wasn't sure if the former was just his accent, and the latter from the lack of articles in Russian.
posted by phliar at 4:36 PM on October 17, 2012
I've had these questions ever since I heard a Science Friday on phage therapy. The Russian (I think) gentleman pronounced it fahj, and used it like a plural, e.g. "this is what phage can do". I wasn't sure if the former was just his accent, and the latter from the lack of articles in Russian.
posted by phliar at 4:36 PM on October 17, 2012
"1. Do you pronounce it fayj or fahj?"
It depends on the speaker, both are standard. I've noticed that folks from commonwealth countries, and particularly Australia, are more likely to say fahj while just about everyone else says fayj. The Dutch and Flemish say faag.
"2. Is phage singular or plural?"
Phage is short for bacteriophage, from the original name given to them in French by D'Herelle. Generally phage is considered to be correct as either singular or plural unless you are referring to a multiple of distinct phage particles - at which point the multiple particles are referred to as phages. If you think thats weird just look a the difference between virus, viruses, virii, virion, and virions.
Incidentally that guy was Sandro (Alexander) Sulakvelidze who is from the Republic of Georgia.
posted by Blasdelb at 5:36 PM on October 17, 2012
It depends on the speaker, both are standard. I've noticed that folks from commonwealth countries, and particularly Australia, are more likely to say fahj while just about everyone else says fayj. The Dutch and Flemish say faag.
"2. Is phage singular or plural?"
Phage is short for bacteriophage, from the original name given to them in French by D'Herelle. Generally phage is considered to be correct as either singular or plural unless you are referring to a multiple of distinct phage particles - at which point the multiple particles are referred to as phages. If you think thats weird just look a the difference between virus, viruses, virii, virion, and virions.
Incidentally that guy was Sandro (Alexander) Sulakvelidze who is from the Republic of Georgia.
posted by Blasdelb at 5:36 PM on October 17, 2012
This is badass. Thanks for the FPP and subsequent comments, Blasdelb.
posted by Sleeper at 8:28 PM on October 17, 2012 [1 favorite]
posted by Sleeper at 8:28 PM on October 17, 2012 [1 favorite]
I've finally processed this.
Nature is bad-ass. The ultimate battlefield. That creatures as complex as us have managed to hold together a semblance of self and species-ness in the face of relentless invasion and destruction from cosmic rays, heavy metals and other toxins, oxidation and other chemical attacks, plasmids, viruses, bacteria, amoebae, and then up to multi-cellular parasites, and then things that feed on us and eat us and use us as transport mechanisms.... it's Sparta, but for reals reals.
And I have figured out why we die! Why everything dies.
It is just the same reason why every two years you should format your PC hard drive. Because after a certain amount of time the cruft and breakage and wastage and disease builds up to such a point that the original functionality is damaged to the point where the best use of the resources embodied is to simply break it up into its smallest components and rebuild it from molecules once again.
Which means, dear friends, that in the end we are all scratch monkeys.
posted by seanmpuckett at 4:44 AM on October 18, 2012
Nature is bad-ass. The ultimate battlefield. That creatures as complex as us have managed to hold together a semblance of self and species-ness in the face of relentless invasion and destruction from cosmic rays, heavy metals and other toxins, oxidation and other chemical attacks, plasmids, viruses, bacteria, amoebae, and then up to multi-cellular parasites, and then things that feed on us and eat us and use us as transport mechanisms.... it's Sparta, but for reals reals.
And I have figured out why we die! Why everything dies.
It is just the same reason why every two years you should format your PC hard drive. Because after a certain amount of time the cruft and breakage and wastage and disease builds up to such a point that the original functionality is damaged to the point where the best use of the resources embodied is to simply break it up into its smallest components and rebuild it from molecules once again.
Which means, dear friends, that in the end we are all scratch monkeys.
posted by seanmpuckett at 4:44 AM on October 18, 2012
So to sum up: Her eyeball had an amoebic infection, which had a giant virus infection, which had a lysogenic virophage infection, which had a transpoviron infection.
I desperately want one of these articles to end with, "Unconfirmed reports indicate that the patient then attempted to remediate her condition by swallowing a fly."
posted by eritain at 6:42 AM on October 19, 2012 [1 favorite]
I desperately want one of these articles to end with, "Unconfirmed reports indicate that the patient then attempted to remediate her condition by swallowing a fly."
posted by eritain at 6:42 AM on October 19, 2012 [1 favorite]
« Older It's Time For Another Woman Moderator for the U.S.... | What do you mean you've never been to Alpha... Newer »
This thread has been archived and is closed to new comments
posted by feckless at 3:59 PM on October 16, 2012 [2 favorites]